|
MEK2-SIPK/WIPK Cascade of Nicotiana benthamiana Is Involved in the Failure of Infection by classd1 Mutant of Colletotrichum lagenarium. Shigeyuki Tanaka1, Nobuaki Ishihama2, Hirofumi Yoshioka2, Gento Tsuji1, Seiji Tsuge1 and Yasuyuki Kubo1 1Laboratory of Plant Pathology, Graduate School of Agriculture, Kyoto Prefectural
University, Kyoto. |
|
| 私はこれまでに、ウリ科植物に病気を起こすウリ類炭疽病菌という植物病原糸状菌の病原性機構について分子生物学的手法を用いた解明を試み、病原性遺伝子ClaSSD1を単離・同定しています。この結果については、前回同窓会の援助を得て参加した国際学会XII International congress on molecular plant-microbe interactionsでも発表しており、また論文として公表されています(以下参照)。 Tanaka S, Yamada K, Yabumoto K, Fujii S, Huser A, Tsuji G, Koga H, Dohi K, Mori M, Shiraishi T, O'Connell R, Kubo Y. Saccharomyces cerevisiae SSD1 orthologues are essential for host infection by the ascomycete plant pathogens Colletotrichum lagenarium and Magnaporthe grisea. Mol Microbiol. 2007 Jun;64(5):1332-49.PMID: 17542924 今回の学会で発表した成果は、この論文からの発展になります。 |
|
|
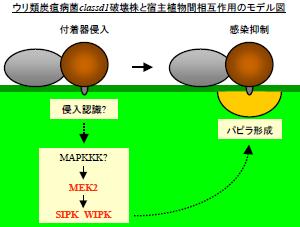
前回の研究成果から、ClaSSD1遺伝子を破壊した変異株は宿主植物に対して病気を起こすことが出来ず、またその原因は変異株の植物に対する感染行動が植物防御応答(パピラ形成)により抑制されることに起因することを細胞学的観察により明らかにしていました。そして次に明らかにすべきはこの植物防御応答因子の解明ですが、そのためには分子レベルでの解明が必須のアプローチとなります。しかし、ウリ類炭疽病菌の宿主であるキュウリでそれを行うのは容易ではありません。そこで、ウリ類炭疽病菌が感染可能であり、なおかつウイルス誘導型遺伝子サイレンシング(Virus-induced
gene silencing; VIGS)が確立されているNicotiana benthamiana(ベンサミアーナタバコ)を植物として用い、classd1破壊株の感染抑制に関与する防御応答因子の探索を試みました。classd1破壊株をN. benthamianaに接種すると、キュウリに接種した場合と同じく病斑は観察されず、パピラ形成による感染行動の抑制が観察されました。そこで、既知の防御応答関連遺伝子である、MEK1, MEK2(MAPキナーゼキナーゼ)、SIPK, WIPK(MAPキナーゼ)、NbrbohA, NbrbohB(活性酸素生成)、RAR1, SGT1, HSP90(R遺伝子型抵抗性)、Syp121/PEN1(非宿主抵抗性)についてVIGSを行い、サイレンシング植物に対してclassd1破壊株を接種し病原性が見られるかどうかを調べました。すると、MEK2とSIPK/WIPKのサイレンシング植物に対して病斑を形成することがわかりました。これらはカスケードを構成していることが知られており、すなわちMEK2-SIPK/WIPKカスケードがclassd1破壊株の感染抑制に関与する植物防御応答因子であることが明らかになりました。このMAPKカスケードをサイレンシングした植物では、ウリ類炭疽病菌野生株に対しても罹病性が高くなります。すなわち、親和性関係における植物の基本的抵抗性を担う因子としての役割を果たしていると考えられます。 classd1破壊株はタマネギ表皮上において野生株より早くパピラ形成を誘導することを明らかにしています。このことから、破壊株は植物防御応答を誘導しているのではないかという仮説を立てています。既にこの仮説を肯定する結果を得ており、今後さらなる検証によりこの微生物と植物の相互作用について新たな知見を得たいと考えています。 |
 |
発表したポスター写真 |
|
| Topへ戻る | |
{kind=link}